日本鳗是一种很珍贵的食用鱼类,生活史非常奇特。属于降海洄游姓鱼类,其洄游的方向与溯河回游性的的鲑鱼相反,在河川中生长,成熟之后洄游到深洋产卵。一生只产卵一次,产后随即死亡。日本鳗的产卵场,根据最新的研究,是位于万里并纳海沟附近的海山产卵(Tsukamoto 2006)。孵化后的仔鱼,外型酷似柳针,故称之为柳叶鱼(Leptocephalus,原义是挟首的意思)。仔鱼随北赤道洋流往西飘送,到了菲律宾周边海域之后进入黑潮流域,继续被飘送到台湾、中国大陆、韩国及日本的陆棚之后,变态为玻璃鳗(Glass eel)。进入河川的入海口时,身上出现黑色素,称之为鳗线(Elvers)。在河川的成长期,称之为黄鳗(Yellow eel)。成熟之后经历第二次变态,眼睛变大,体色变黑,肠道萎缩,卵巢持续发育,称之为银鳗(Sliver eel)。至此不在进食,开始游向产卵场,准备产卵。
鳗鱼一生充满传奇性故事,全世界总共有18种鳗鱼,分别分布于印度洋、太平洋、大洋洲及大西洋的美州及欧洲 (Tesch 1977),生活史模式都很类似。首先揭开鳗鱼生活史谜团的,是丹麦的科学家史密特博士(Schmidt 1922),经过长期的追踪才发现欧洲鳗在藻海(Sargasso Sea)产卵。日本于50年代即开始寻找日本鳗的产卵场(Matsui 1957),直到最近才发现其真正的产卵场位置(Tsukamoto 2006)。50多年来,花了很多的时间,主要原因是假设错误及生活史知识不足。近20年来,由于研究工具的创新以及调查频率增加,鳗鱼生活史的研究有了很大的进展。因长期捕捞,世界上几种主要的鳗鱼种类(包括美洲鳗、欧洲鳗及日本鳗),其资源量已经下降到1980年代资源水平的1?10%(Dekker 2003),虽然不至于立刻绝种,但已经到了濒危边缘,资源下降的原因不是很明显,但其原因不外乎全球气候变迁,河川据地恶化和污染,寄生虫感染以及过度捕捞等因素。尤其是每年在河口捕捞,用来养殖的鳗线之数量非常惊人,造成河川及内陆湖泊的鳗鱼数量急遽减少。为了生产人工鳗苗以减少天然鳗苗的捕获压力,近年来的鳗鱼人工催熟技术及育苗技术,已经有很大突破。
一、引言
黑潮是西北太平洋的海水输送大动脉。黑潮是源自于北赤道洋流的一股暖流。因为黑潮的流经,台湾的冬天,比起同纬度的邻近地区温暖许多。每年冬季,成千上万的鳗线顺着寒流涌向河口地区,沿岸渔民下网捕捞,卖给鳗鱼养殖户,可带来一笔可观的财富。鳗鱼是台湾很重要的养殖鱼类,是日本料理中的一道美食。但是很少人知道,鳗线在小时候是乘着黑潮暖流而来的。本文将从鳗鱼的生活史开始,介绍黑潮与鳗线之关系以及鳗线产量的变动。
二、鳗鱼的生活史
全世界的鳗鱼有十八种及亚种 (Tesch 2003),除了欧洲鳗 (Anguilla anguilla) 及美洲鳗 (A. rostrata) 分布在北大西洋之外,其余16种皆分布在印度太平洋一带 (图一)。鳗鱼的祖先种最先出现在印度尼西亚海域。根据DNA的考证,两种大西洋鳗大约在一亿年前经由古赤道海或五万年前由巴拿马海峡进入大西洋的 (Aoyama et al. 2001, Lin et al. 2001)。台湾地区共发现四种鳗鱼,分别为日本鳗 (A. japonica)、鲈鳗 (A. marmorata)、西里伯斯鳗 (A. celebesensis) 及短鳍鳗 (A. bicolor pacifica) (Tzeng 1982, Tzeng and Tabeta 1983)。日本鳗为温带鳗,其它三种为热带鳗。鲈鳗会上溯到高山溪流,可生长至1公尺以上 (图二)。近年因栖地破坏,野生的鲈鳗非常少,政府早就将之列为保育类动物,禁止捕捞。西里伯斯鳗及短鳍鳗为南洋品种,台湾位于其分布边缘,偶而会出现。日本鳗是四种中产量最多的,近年来其野生数量日渐减少,市面上的日本鳗几乎都是养殖的 (Tzeng 2004)。
日本鳗与其它鳗鱼一样都是属于两侧洄游性鱼类,在河川中长大,成熟之后奋力洄游到海洋中产卵。日本鳗每年秋冬季降海,大约半年的时间洄游到产卵场,一生只产一次卵,就结束生命。鳗鱼的生活史可分为卵期 (Egg-stage)、柳叶鱼期 (Leptocephalus)、玻璃鳗期 (Glass eel)、鳗线期 (Elver)、黄鳗 (Yellow eel) 及银鳗 (Silver eel) 等六个时期 (图三)。日本鳗的产卵场位于太平洋最深处的马里亚纳海沟西方海域 (Tsukamoto 1992, 2006)。孵化之后的柳叶鱼,顺着北赤道洋流往西漂,到了菲律宾东方海域之后,进入黑潮往北漂送,途经台湾、中国大陆、韩国及日本的大陆棚水域,然后由沿岸水送到了河口域,这时身体表面出现黑色素,称之为鳗线。黑潮在柳叶鳗的分布上扮演非常重要的角色,而鳗线则在沿岸水的推波助澜之下,进入河口域哺育。日本鳗鳗线从产卵场到达河口域,大约需要半年的时间 (Cheng and Tzeng 1996)。鳗线进入河川中的成长阶段,称之为黄鳗。黄鳗经过五至八年的成长之后,身体成为银灰色,性腺开始成熟、眼睛变大、肠道萎缩,准备降海产卵,这时候的鳗鱼称之为银鳗 (Tzeng et al. 2000, Han et al. 2003)。
三、产卵场的发现
日本鳗的产卵场,已知是在北赤道洋流上的马里亚纳海沟的西方海域 (图四)。产卵场的发现是经过一段非常漫长岁月的调查才确认的。最早松井魁博士认为日本鳗的产卵场是在台湾东方海域 (Matsui 1957)。他会做这样的推测的理由是因为,日本鳗在陆地上的分布最南限为台湾。如果日本鳗就近产卵的话,产卵场应该就在台湾东方海域。后来郭河先生根据台湾西部沿岸的鳗线比东部多的情况来看,认为台湾西南海域应该另有一个产卵场,否则无法解释台湾沿岸鳗线的分布状况,因为鳗线若顺着黑潮往北漂的话,只有台湾东部沿岸会出现鳗线 (Kuo 1971)。但是经过三十年的摸索,以上两个假说却都无法得到证实。之后从鳗线耳石日周轮研究 (Tabeta et al. 1987) 以及黑潮的流速来看,鳗鱼的产卵场应该要比上述两个海域更远才对,于是在1991年及最近 (2005年) 的大规模调查才发现,鳗鱼真正的产卵场应该在马里亚纳岛西侧 (Tsukamoto 2006)。
四、耳石的微细构造所记录的鳗鱼的洄游过程
鱼类的耳石有3对,分别为矢状石 (Sagitta)、扁平石 (Lapillus) 以及星状石 (Asteriscus)。耳石的主要生理功能,为感觉及平衡。耳石会随着鱼体的日夜韵律及季节性成长而同步增大,耳石经过适当处理之后会出现日周轮及年轮,可以用来判读鱼类的日龄及年龄。耳石是一种生物矿化的结晶,主要成分为碳酸钙 (CaCO3)。环境中的微量元素会经由鱼体的吸收进入耳石中。由耳石中的元素变化,可以重建鱼类的洄游环境史。因此,耳石是时间和环境条件的记录器 (图五)。
图六是1995年8月22日台湾省水产试验所水试一号在产卵场附近所采集的一尾体长28.8 mm的日本鳗柳叶鱼。将其耳石取出,经过研磨及稀酸腐蚀之后,在电子显微镜下呈现出明显的日周轮。该耳石像不像一片光盘片?由日周轮数得知是出生后45天的柳叶鱼,逆算后其生日为7月3日 (亦即农历6月4日),换言之,鳗鱼是在新月期间产卵 (Liao et al. 1996)。
图七是在河口域采获的鳗线的耳石局部放大照片。有人把耳石比喻为罗塞达石碑 (Rosetta stone)。罗塞达石碑是一块很有名的石块,该石块同时刻有埃及象形文字的神圣体、通俗体以及古希腊文三种文字,因为它的发现使得埃及象形文字得以推考。商博良 (J.-F. Champollion, 1790~1832),因为这项发现而被尊称为埃及文之父。同样的道理,耳石上刻有鳗鱼生活史的轨迹,借着耳石的解读,我们也可以了解鳗鱼的生活履历。从耳石的微细构造 (Microstructure),我们发现每一个发育阶段的耳石构造的纹路不同。从鳗线的耳石上可以找到几个明显的地标点,分别为孵化轮 (HC)、第一次摄饵轮 (FFC)、变态轮 (MC) 及鳗线轮 (EC),其所对应的发育阶段分别为胚胎期 (Embryo)、卵黄囊期 (Yolk-sac Stage)、柳叶鱼期 (Leptocephalus)、玻璃鳗期 (Glass eel) 及鳗线期 (Elver)。由日周轮的计算,可以知道每一个阶段的持续时间,以及回推鳗鱼的生日和柳叶鱼变态时的日龄 (Cheng and Tzeng 1996)。
五、耳石微化学所记录的鳗鱼的生活史事件
耳石的化学组成中,除了钙 (Ca) 之外,还有一种含量很高的元素,锶 (Sr)。海水中锶的浓度是淡水的一百倍。随着外在锶浓度的改变,耳石中锶的浓度也会有明显的变化。鳗鱼从柳叶鱼变态为玻璃鳗之后,接着就要从海水游向淡水。因此,由耳石锶浓度的变化就可以回推其变态的时间点。图八是鳗鱼由产卵场游向河口域的过程中,耳石所记录的锶钙比 (Sr/Ca ratio) 的变化 (Tzeng and Tsai 1994)。将图八和图七比对,就可以找出鳗线耳石上柳叶鱼变态的痕迹 (MC)。耳石锶钙比的测定需要精密的仪器,电子微探仪 (EPMA, Electron Probe Microanalyzer)。
六、由耳石所纪录的事件来还原鳗线的生活履历
柳叶鱼及玻璃鳗的空间分布明显不同 (Tzeng 2003)。从图四得知,不同发育阶段的个体,其分布是泾渭分明的,柳叶鳗由黑潮输送到东北亚陆棚后,即变态成为玻璃鳗,然后顺着沿岸水进入河口域。换言之,随着发育阶段的变化,必须转移栖息环境,否则无法从产卵场游到河口进入河川中生长。变态的速率若差之毫里,到达的目的地则失之千里。
表一是我们从日本鳗鳗线分布的几个沿岸国家 (包括台湾、中国大陆及日本) 的河口域所采集到的鳗线 (图四)。根据每尾鳗线的耳石的微细构造及微化学变化,定出上述发育阶段的地标点,并计算变态日龄 (Tm) 及变态之后至到达河口的时间,亦即玻璃鳗的持续时间 (Tr-m)。结果发现,当变态日龄延后时,其到达的国家或河口就不同。例如,Tm从115.8 (S2) 增加到137.0 (I),也就是延迟21天变态时,柳叶鱼就被黑潮从台湾带到日本。也就是说,以其延迟变态的时间 (∆t) 乘上黑潮的流速 (υ),足足可以使柳叶鱼在海上多漂流2016公里 (Cheng and Tzeng 1996)。
七、鳗线的漂送与沿岸流的关系
柳叶鱼变态之后,便脱离黑潮流域,因此其洄游路径,主要是受沿岸流的影响 (图九) (Tzeng 1997)。从耳石的纪录来看,台湾南部的鳗线的玻璃鳗期 (Tr-m) 比北部长 (表一),这证明台湾南部的鳗线是从北往南输送的。也就是说,过去为了以黑潮的输送来解释台湾西部沿岸鳗线较多的原因,而假设台湾西南海域有一个产卵场是不正确的 (Kuo 1971)。因为台湾西岸的鳗线是顺着中国大陆沿岸流南下的,而不是顺着黑潮北上的。
八、鳗线渔获量的变化
台湾的养鳗业从1965年开始产业化,因此每年鳗苗 (线) 的需求量不断加,但是天然鳗线的捕获量并没有跟着增加,而是呈现周期性的变动 (图十)。最高的年捕获量还不到两亿尾,但最大的年需求量却超过五亿尾。不足的部分需仰赖进口 (Tzeng 1997)。
鳗线来游量的变化,可分为一个渔期内的日变化,与长周期的年变化。日变化与月亮周期、潮汐及水温有关 (图十一)。沿岸冷水最强 (水温最低) 时,来游量到达最高峰,这证明上述鳗线是随着中国大陆沿岸冷水洄游而来的假说。另外鳗线对光线非常敏感,一个月两次大潮中,只有新月才出现高峰,满月时则受到光线抑制而不活跃,渔获量不振 (Tzeng 1985)。另外,根据鳗线的年产量资料,以周期性解析法 (Periodogram) 分析结果,发现鳗线的来游呈现大约十年的周期变化 (Tzeng 1997)。自然界的十年周期为太阳黑子的活动周期,鳗鱼十年周期的渔获量变化,究竟是什么原因,目前还不清楚。
日本鳗和大西洋的欧洲鳗及美洲鳗一样,1980年以后呈现急速下降的趋势(图十二),同时也发现鳗线渔获量的变化与全球暖化及圣婴现象有关,在圣婴年时,鳗线产量有降低的现象,不论是台湾、中国大陆及日本皆然(图十三至十五)。长期来看,日本及台湾的鳗线产量呈现下降趋势,但中国大陆则呈现增加的趋势,这可能与近年来中国投入捕获的人力(渔获努力量)的增加有关。
九、结语
鳗线从产卵场洄游到河口域,花费长达半年的时间,而且历经一连串的发育阶段变化,以及不同的海洋环境。若要还原这些过程,必须耗费大量的人力、物力以及长时间的海上采集,才能拼凑出其发育阶段的变化及洄游过程。但是耳石忠实地记录了鳗鱼生活史的细节,从耳石的分段解析来看,从产卵场到河口域,柳叶鱼主要是受到北赤道洋流和黑潮的影响,变态之后的鳗线主要是受沿岸流影响。耳石内尚蕴藏很多有趣及神秘的生活史讯息,还有待进一步的发掘。
目前鳗鱼人工繁殖技术,可以从卵孵化阶段养到黄鳗阶段,但柳叶鱼需耗时200天才会变态,比起天然的100天左右要长很多,而且成功的例子很少,仅此于实验阶段,还不能商业化。养殖的鳗苗,全赖天然捕捉,产量又不安定,而且欧盟已经开始实施鳗苗输出的管制。鳗苗的来源是养鳗业成功与否的关键,其产量的变化,值得业者重视。
.jpg)
图 1. 鳗鱼的世界分布 (Tesch 2003)
.jpg)
图 2. 鲈鳗 (Williamson 1993)
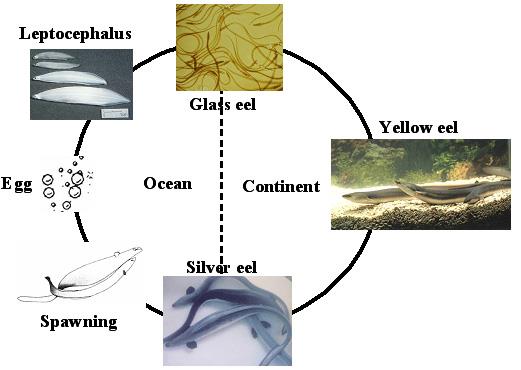
图 3. 鳗鱼的两侧洄游生活史
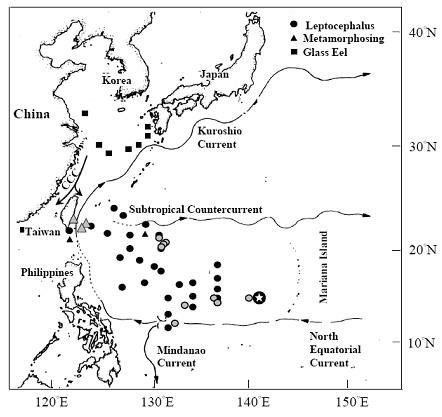
图 4. 日本鳗产卵场( )、柳叶鱼及玻璃鳗的分布与洋流之关系 (Tzeng 2003)
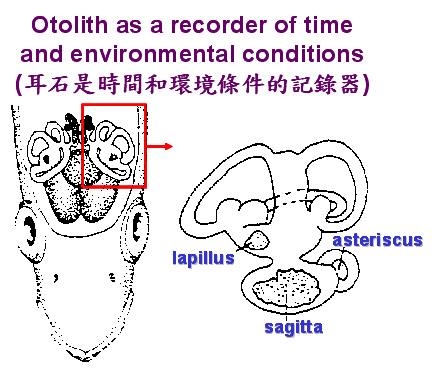
图 5. 真骨鱼类三对耳石的相对位置
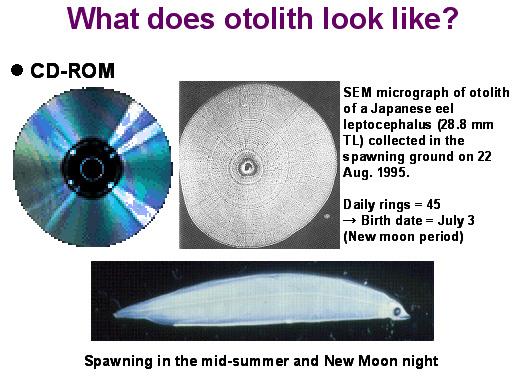
图 6. 日本鳗的柳叶鱼及其耳石
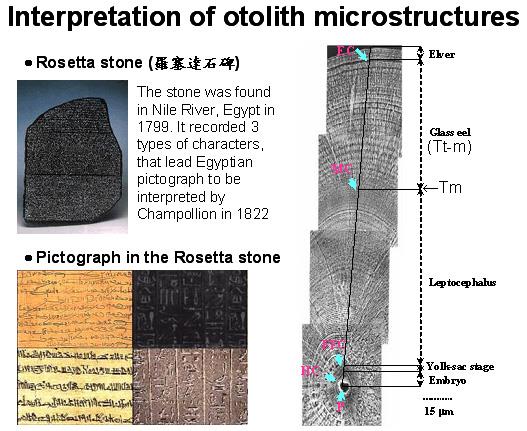
图 7. 日本鳗鳗线耳石微细构造与发育阶段之对应关系
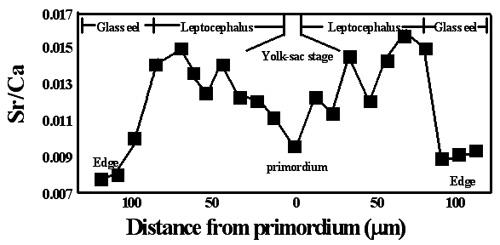
图 8. 日本鳗鳗线耳石锶钙比与发育阶段之对应关系 (Tzeng and Tsai 1994)
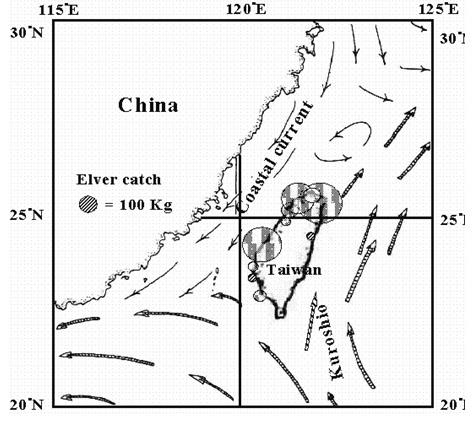
图 9. 日本鳗鳗线的分布与沿岸流之关系 (Tzeng 1997)
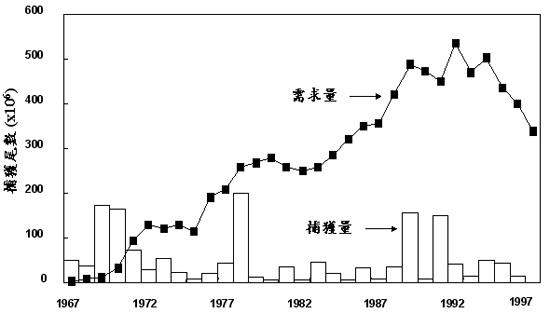
图 10. 历年日本鳗鳗线的捕获量与需求量 (Tzeng 1997)
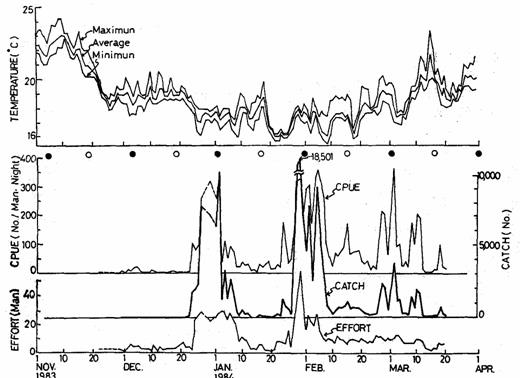
图 11. 日本鳗鳗线来游量的变化及其与水温和月亮周期之关系(Tzeng 1985)
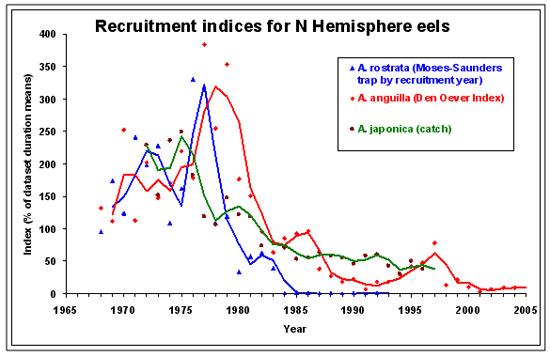
图 12. 日本鳗、欧洲鳗及美洲鳗的变动趋势(仿自Knights 2006 )
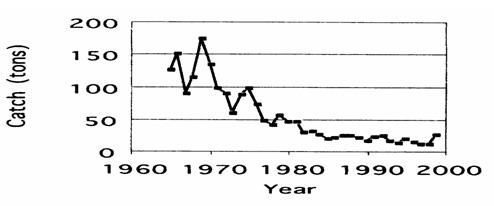
图 13. 日本的鳗线年产量(仿自Tatsukawa 2001)
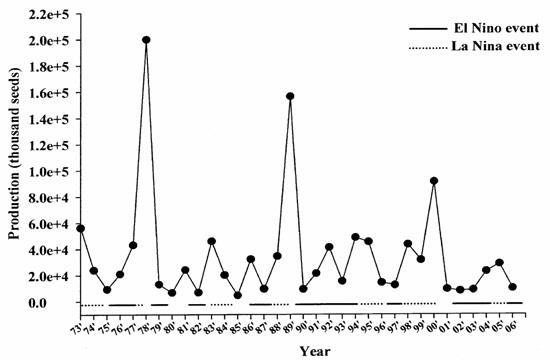
图 14. 台湾鳗线产量的年变化与圣婴现象的关系(Han et al. 2007未发表)
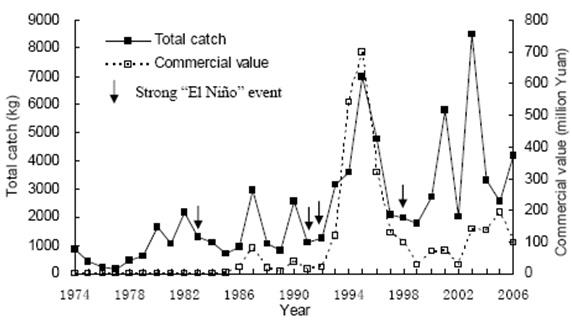
图 15. 扬子江口鳗线产量及产值的年变化之关系与圣婴现象的关系(仿自Zhang et al.2007)。资料来源Shanghai City authority)
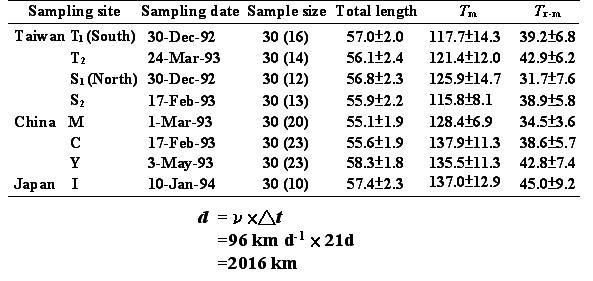
图 16. 表一、日本鳗柳叶鱼变态日龄 (Tm) 及玻璃鳗持续时间 (Tr-m) 的地区间比较 (Cheng and Tzeng 1996)

中国鳗鱼网(www.chinaeel.cn)所刊登的所有内容,包括但不限于图片、文字及多媒体形式的新闻、信息等,未经著作权人合法授权,禁止一切形式的下载、转载使用或者建立镜像。获得合法授权的,应在授权范围内使用,必须为作者署名并注明“来源:中国鳗鱼网”字样。违反上述声明者,本网将依法追究其相关法律责任。
